
Estudio de la tasa fotosintética en relación a la concentración de pigmentos fotosintéticos en plantas de hojas de coloración verde y hojas rojo-granate
Inés Valencia Fernández
Resumen
La fotosíntesis, realizada por los organismos autótrofos, es el único proceso biológico capaz de aprovechar la energía lumínica del Sol gracias a la acción de los pigmentos fotosintéticos. El desarrollo de las plantas depende de este proceso, pudiéndose observar cómo las plantas de hoja verde han alcanzado un mayor grado de diversificación que las plantas de hoja roja-granate, ¿esto determina que existe una relación entre los pigmentos fotosintéticos y la tasa de fotosíntesis?
En esta investigación se estudiaron diez plantas distribuidas en un grupo de hojas verdes y un grupo de hojas rojas-granates. En primer lugar se realizaron cuatro cromatografías a cada una de ellas para conocer su cantidad media de pigmentos fotosintéticos. Posteriormente, se dispuso un montaje especialmente preparado para esta investigación con el que mantener constantes las condiciones de las plantas, para monitorizar el proceso de fotosíntesis. Las plantas estaban aisladas de variables que pudieran interferir negativamente en el proceso, pero podía medirse con sensores específicos la cantidad de oxígeno producido en el recinto cerrado en el que se encontraban.
Analizados los resultados de diferenciación de pigmentos fotosintéticos y medición de tasas de fotosíntesis, se ha concluido que la coloración externa de las plantas se corresponde con el pigmento fotosintético mayoritario que presentan, pero ello no influye en su tasa de fotosíntesis, puesto que no hay diferencias significativas entre las tasas registradas en plantas de hoja verde y plantas de hoja roja-granate. Estas últimas, con mayor cantidad de carotenos, llegan a igualar las tasas de las plantas de hoja verde, aunque presentan más clorofila, pigmento principal en la fotosíntesis. Por tanto, puede establecerse que la distribución pigmentaria y la tasa de fotosíntesis de las plantas angiospermas, son dos variables que no se ajustan a ninguna proporción, sino que se adaptan a las necesidades de las distintas especies.
Agradecimientos
Sin duda, esta investigación es el reto más amplio al que me he enfrentado durante el curso de Bachillerato Internacional. Igualmente, es el proyecto en el que más activamente he trabajado y con el que he podido terminar de comprobar la cantidad de apoyos con los que he contado en estos dos años, en los que no todo han sido buenos momentos.
Por esto mismo quiero agradecer a mis padres, Manuel Valencia y Jerónima Fernández, el ánimo que me han dado y la confianza que han depositado en mí no sólo en este tiempo, sino en todo momento. Tampoco puedo olvidarme de mi tutor de monografía, Juan García, y de mi profesora de biología, Aurora Aparicio, por ayudarme a aclarar mis ideas en el comienzo de esta experimentación y por hacer crecer en mí un interés inmenso por todo lo que esta ciencia arrastra.
Confío en que esta investigación signifique tan sólo el comienzo de otras que vendrán y, por qué no, que siempre quede un hueco abierto para retomarla algún día y recordar lo mucho que he aprendido con ella.
1. Introducción
La fotosíntesis puede considerarse, desde un punto de vista biológico, el principal proceso de metabolismo anabólico desarrollado en la naturaleza, ya que la gran mayoría de los seres vivos tienen una dependencia directa de los productos sintetizados en él. A grandes rasgos, la fotosíntesis consiste en la producción de materia orgánica y la liberación de oxígeno a partir de agua y otras sustancias inorgánicas. Sin embargo, al estudiarla de manera global, puede observarse cómo la importancia de la fotosíntesis radica en otro suceso: la transformación de la energía lumínica del Sol en energía química asimilable por los seres vivos. Por este motivo la energía lumínica es el elemento primordial de la fotosíntesis, ya que sirve como combustible para desarrollar sus diferentes reacciones metabólicas. Sin embargo, a pesar de su cuantioso valor, es el elemento más complejo de captar y asimilar por los organismos autótrofos. Por esto mismo los organismos autótrofos constan de unas moléculas específicamente encargadas de absorber la luz del Sol y hacer que participe activamente en la fotosíntesis: los pigmentos fotosintéticos.
Las hojas de las plantas contienen gran variedad de pigmentos, como así muestra la diversidad de colores que presentan, sin embargo, pocos de estos son realmente pigmentos fotosintéticos. Por tanto, ¿puede asegurarse que el color que presentan las plantas se corresponde con su cantidad de pigmentos fotosintéticos? Si así fuera, el hecho de que las plantas de hoja verde sean las más abundantes y se hayan adaptado a tantos ecosistemas, indicaría que han desarrollado algún tipo de ventaja adaptativa, es decir, una mayor eficacia en el proceso de fotosíntesis en comparación con las plantas de otra coloración, por ejemplo, las plantas con hojas rojas-granate.
De este planteamiento surgió el fondo de esta investigación: ¿existe relación entre los pigmentos fotosintéticos de una planta y su tasa de fotosíntesis? En las páginas siguientes se realizará un estudio y comparación entre la cantidad de pigmentos fotosintéticos y la tasa de fotosíntesis de diez especies diferentes de plantas, clasificadas en plantas de hoja verde y plantas de hoja roja-granate:
| Plantas de hoja verde | Plantas de hoja roja-granate |
|---|---|
| Chlorophytum comosum | Begonia rex |
| Hedera helixomosum | Coleus blumei |
| Impatiens wallwriana | Iresine herbistii |
| Piectranthus australis | Tradesencantia andersaniana roja |
| Syngonium podophyllum | |
| Tradesencantia andersaniana verde |
2. Generalidades
2.1. Fisiología de la fotosíntesis
La mayoría de los organismos que desarrollan la fotosíntesis conforman la base de las cadenas tróficas que se desarrollan en los diferentes ecosistemas. Esta investigación se centrará en el estudio de las plantas angiospermas, por ser los organismos autótrofos con los que los metazoos tenemos más relación, debido la energía química que recibimos de ellos.
La mayoría de las células vegetales que componen una planta presentan orgánulos específicos en los que se desarrolla el proceso de fotosíntesis: los cloroplastos. El tejido del mesófilo, en la parte interior de las hojas, es el que presenta las células más ricas en cloroplastos (30-40 por célula); por esto mismo esta investigación está centrada en el estudio de las hojas de las plantas. Concretamente, dentro de los cloroplastos aparece un líquido denso denominado estroma en el que, pueden diferenciarse unos sacos membranosos llamados tilacoides, apilados verticalmente formando la grana.
Los tilacoides están rodeados por una bicapa lipídica que cumple un papel fundamental en las primeras etapas de almacenamiento de energía en el proceso de fotosíntesis1. Es en este lugar donde se encuentran los pigmentos fotosintéticos asociados a proteínas de membrana. Definitivamente, éste es el lugar preciso en el que se desarrollarán las reacciones metabólicas de la fotosíntesis, comenzando por la acción fotoreceptora de los pigmentos fotosintéticos explicada a continuación.
2.2. Características de la luz solar
La fotosíntesis consta de dos etapas diferenciadas: la fase dependiente de la luz y la fase independiente de ésta. Observándose así la importancia de la primera fase como promotora del proceso, fruto de la acción de los pigmentos fotosintéticos. Por tanto, para conocer la actuación de éstos, deben conocerse previamente las características básicas de la luz solar.
La luz emitida por el Sol es una onda electromagnética. Si bien el espectro de radiación electromagnética engloba longitudes de onda desde menos de un nanómetro (rayos gamma) hasta más de un kilómetro (ondas de radio), la luz aprovechable por los organismos autótrofos es la angosta franja que abarca entre los 380 nm y los 750 nm2.
A pesar de que la luz se propaga atendiendo a sus propiedades ondulatorias, la condición indispensable para comprender su intervención en procesos metabólicos, es que debe entenderse como un complejo de partículas, denominadas fotones. Los fotones tienen una energía asociada, estableciéndose una relación de proporcionalidad inversa con respecto a la longitud de onda que posee el rayo lumínico del que provienen. De modo que un rayo de luz con menor longitud de onda presenta mayor energía.
Respecto a este postulado, el fisiólogo y botánico alemán Theodor Wilhelm Engelmann3 desarrolló en 1883 una investigación con la cual determinó que los rayos de luz con una longitud de onda entre 450 nm y 670 nm (correspondiente a las secciones azul y roja del espectro), permitían la máxima eficacia de la fotosíntesis. Estas longitudes de onda se asemejan al rango de absorción del pigmento fotosintético de la clorofila, causa por la que se le considera el pigmento más activo en la fotosíntesis.
En cambio, al comparar la representación del espectro de acción de la fotosíntesis obtenido en el experimento de Engelmann con el espectro de absorción de la clorofila, puede verse cómo ambas distribuciones no se ajustan perfectamente. De aquí puede deducirse que la clorofila no actúa sola en el proceso de fotosíntesis, sino que su combinación con otros pigmentos accesorios es lo que permite que en el proceso se alcancen niveles de energía elevados.

2.3. Pigmentos fotosintéticos
Cada pigmento fotosintético presenta mayor afinidad respecto a una longitud de onda dentro del espectro de la luz visible; absorben unas longitudes de onda determinadas del espectro y, con ello, su energía asociada.
Los pigmentos fotosintéticos corresponden a una misma composición química: son tetraterpenos, derivados del isopreno 2-metil-1,3-butadieno. En su estructura molecular comparten una sección denominada fitol, una larga cadena hidrocarbonada con restos de metilo que presenta enlaces dobles conjugados, de los que son liberados los electrones que participan en la fase lumínica de la fotosíntesis.
Las características específicas de los pigmentos estudiados en esta investigación son:
- Clorofilas: la molécula de clorofila está formada por cuatro anillos de pirrol entrelazados que forman un anillo más general, la porfirina, en cuyo interior aparece un átomo de Magnesio (Mg2+). Existen sendos tipos de clorofilas con pequeñas fracciones estructurales, sin embargo, en la investigación que prosigue no se hará diferenciación entre ellas.
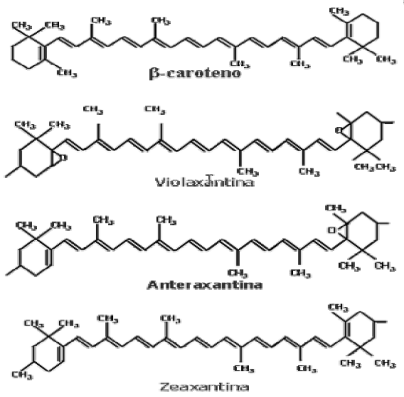
- Carotenoides: dentro de este grupo están englobados carotenos y xantofilas. Su estructura básica es lineal e hidrocarbonada, con la sucesión de más de 40 carbonos. La diferencia entre estos dos pigmentos radica en que los carotenos únicamente presentan carbono e hidrógeno, mientras que las xantofilas poseen oxígeno en sus anillos terminales6.
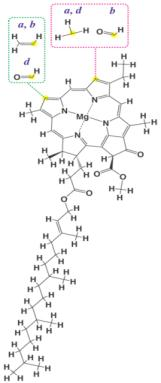
2.4. Transformación de la energía lumínica en energía química
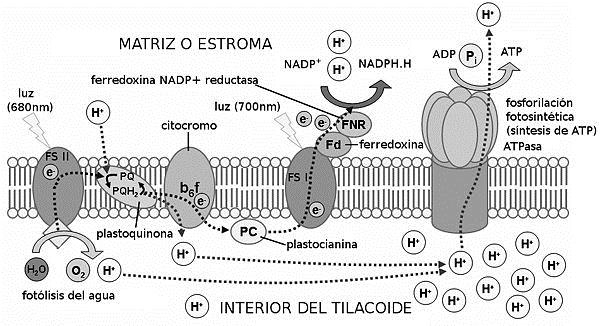
Los pigmentos fotosintéticos se organizan formando fotosistemas, mecanismo compuesto por dos estructuras: centros de reacción (complejo proteico formado por dos moléculas de clorofila) y complejos antena (moléculas de pigmentos accesorios ligados a proteínas de membrana). Se diferencian dos fotosistemas en función al rango de las clorofilas que los componen: PSI con absorción en los 700 nm y PSII en los 680 nm.
El proceso comienza con una primera etapa en la que los complejos antena asociados al PSII captan los fotones de la luz solar y expiden su energía al centro de reacción al que están asociados. Una de las moléculas de clorofila se oxida al liberar un electrón, excitado hasta un nivel superior de energía. Dicho electrón es recogido por la plastoquinona, una sustancia aceptora de electrones que inicia la transferencia hacia distintos factores. Finalmente la proteína plastocianina cede los electrones al PSI y, desde allí, comienza una segunda etapa de trasmisión, promovida por la proteína ferrodoxina.
La acción de la energía lumínica produce la fotólisis del agua, su ionización. De los átomos de hidrógeno liberados se tomarán los electrones que reducirán las moléculas de clorofila de ambos fotosistemas. Por otro lado, el oxígeno será liberado al exterior de la planta; aspecto que se estudiará en esta investigación y que fundamenta el hecho de la posible relación existente entre ambas variables ya que la expulsión de oxígeno deriva de las reacciones iniciadas por los pigmentos fotosintéticos.
Las primeras transformaciones energéticas se producen en la primera etapa, los electrones excitados van perdiendo energía hasta volver a su nivel fundamental. La energía que pierden se utiliza en el proceso de síntesis de ATP (fotofosforilación). Por otro lado, en la segunda etapa de transferencia, los electrones y los protones del agua llegan hasta una molécula de NADP+ que es reducida a NADPH. Finalmente, el ATP y el NADPH otorgarán la energía química y el poder reductor necesarios para la síntesis de carbonatos en la siguiente fase de la fotosíntesis, la fase independiente de la luz, en la que los pigmentos fotosintéticos no participan.
3. Investigación
Esta investigación se ha desarrollado a partir de una mera observación general: la evolución de las plantas de hoja verde ha sido más fructífera que la de las plantas de hoja roja-granate, como muestra su gran diversificación y adaptación a multitud de ecosistemas. La hipótesis de partida establece que las plantas de hoja verde deben presentar algún tipo de ventaja adaptativa sobre las plantas de hoja roja-granate. Como la característica más evidente en la que presentan diferencias es su coloración externa, a continuación se estudiará si existe algún tipo de relación entre la cantidad de pigmentos fotosintéticos y el rendimiento de la tasa de fotosíntesis en diferentes especies.
Se plantearon dos itinerarios paralelos en la investigación: por un lado, el registro de la composición de pigmentos fotosintéticos de cada planta, con objetivo de conocer si la apariencia externa de las hojas de cada especie se corresponde efectivamente con el color del pigmento fotosintético mayoritario que presentan. Y por otro lado, la monitorización del proceso de fotosíntesis para conocer las posibles diferencias entre la tasa de producción de oxígeno de plantas de hoja verde y hoja roja-granate y, analizar si existe dependencia entre éste registro y su distribución pigmentaria.
3.1. Fase de preparación
La primera fase de la investigación consistió en elegir las especies de plantas angiospermas que iban a estudiarse y fijar diferentes constantes para asegurar la igualdad de condiciones en las que se desarrollarían todas ellas. El hecho de que existe mayor diversidad de plantas de hoja verde que de plantas de hoja roja-granate ha podido comprobarse en el desarrollo de la propia monografía, ya que fue más complicado conseguir especies de hoja roja-granate por su escasez. De hecho, se han comparado datos de seis plantas de hoja verde frente a cuatro de hoja roja-granate:
| Plantas de hoja verde | Plantas de hoja roja-granate |
|---|---|
| Chlorophytum comosum | Begonia rex |
| Hedera helixomosum | Coleus blumei |
| Impatiens wallwriana | Iresine herbistii |
| Piectranthus australis | Tradesencantia andersaniana roja |
| Syngonium podophyllum | |
| Tradesencantia andersaniana verde |
La mayoría de las plantas comenzaron a investigarse cuando ya tenían un crecimiento avanzado y un porte considerable, de modo que la fase previa a la toma de datos consistió en estabilizar las variables que podrían influir en el desarrollo del proceso de fotosíntesis: medio de cultivo, exposición a la luz, riego y temperatura.
- Primero se unificó el sustrato en el que crecían las plantas, con tiempo suficiente para que se evidenciaran posibles problemas de éstas frente a las nuevas condiciones. No se adicionó ningún tipo de precursor de crecimiento ni anti-plagas.
- El tiempo de exposición a la luz fue de 6 horas al día, sin establecer ninguna limitación, las plantas no estaban en un invernadero ni cubiertas por un toldo o similar.
- La frecuencia de riego se estableció en una vez cada 24 horas manteniendo el agua con el que las plantas se habían regado en su lugar de procedencia, es decir, las aguas graníticas de Colmenar Viejo (Madrid). Las plantas fueron trasladadas a un segundo lugar durante la investigación, dónde el agua corriente, con alta concentración calcárea, se consideró poco adecuada para las plantas ya que podría afectar a su desarrollo.
- El control de la temperatura no se basó en variar las condiciones para que las plantas siempre estuvieran en un entorno con la misma temperatura fija, lo que se procuró es que en todo momento las plantas se hallaran en el mismo lugar, para que todas estuvieran en un medio con las mismas variaciones de temperatura.
Pasado un mes desde el comienzo de esta fase de preparación, se comenzó con las pruebas de cuantificación de pigmentos fotosintéticos con técnicas cromatográficas y, el registro del proceso de fotosíntesis utilizando sensores específicos, cuyos resultados se analizarán conjuntamente.
3.2. Cromatografías
El fundamento práctico de la separación de pigmentos por cromatografía es bastante sencillo, pero se decidió seguir las indicaciones expuestas en el Manual de Prácticas de Fotosíntesis9, desarrollado por la Facultad de Ciencias de México, ya que ofrecía las especificaciones que mejor se ajustan a la extensión de investigación.
El fundamento de la extracción de pigmentos fotosintéticos por cromatografía se basa en la separación de los pigmentos fotosintéticos por medio de su afinidad con el disolvente apolar con el que se mezclan, ya que son moléculas insolubles en agua. Respecto a lo mencionado anteriormente acerca de la estructura molecular de los pigmentos fotosintéticos, la fracción alifática terminal denominada fitol es lo que les aporta la propiedad de ser insolubles. Por otro lado, los grupos hidroxilo de las xantofilas hacen que sean los pigmentos más solubles en disolventes apolares, seguidos por los carotenos y, por último las clorofilas.
Se realizaron cuatro cromatografías a cada una de las especies. Se efectuaron en días distintos, pero cada uno de los días se realizaba la cromatografía de las 10 especies, para que hubieran estado expuestas a las mismas horas de sol y cantidad de riego. El método se detalla a continuación:
- Se tomaron aproximadamente 10 g de hojas, escogiendo todas ellas de la zona alta de las plantas. Se lavaron y se les retiraron los nervios.
- Se trituraron en un mortero de madera, añadiendo sal para facilitar el majado.
- Se añadieron 40 mL de disolvente extractante, en este caso, alcohol etílico al 90%. El alcohol tiene menor carácter apolar que otros disolventes como el benceno o el éter de petróleo, pero no se disponía de ellos.
- Se colocó la muestra en unos recipientes altos y estrechos (vasos de 6 cm de diámetro y 15 cm de alto); sujetando una tira de papel de filtro de 16 cm de largo y 3 cm de ancho en la parte superior del recipiente, sin dejar que llegara al fondo.
- Se dejó el preparado en un lugar seco durante 24 horas hasta que la marca de absorción de la mezcla cubría por completo el papel de filtro.
- Se dejaron secar las diferentes tiras de papel y, posteriormente se midieron las bandas de cada uno de los pigmentos fotosintéticos con una regla convencional, con un error de 10 mm. Como las bandas de coloración no presentaban mucha diferencia, la precisión de este instrumento no era adecuado para discriminar diferencias en la anchura de las bandas. Debido a esto se repitieron las mediciones con un calibre digital, que ofrece un error de 0,01 mm, mucho más apropiado y que evidenciaba las pequeñas diferencias entre las bandas.
- Como las bandas de diferente color no eran completamente horizontales, se midió desde la zona más alta a la más baja de cada banda. Las bandas correspondientes a las clorofilas a y b se unieron en una misma medición por la dificultad al diferenciar su coloración.

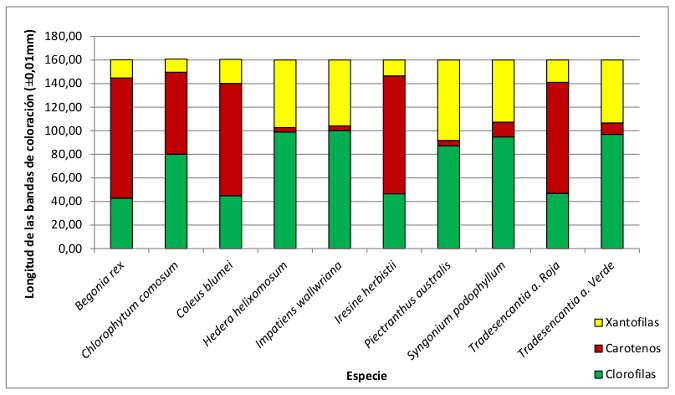
A continuación se presenta la representación de la longitud media de las bandas de coloración de cada una de las especies. Se considera que la longitud de estas bandas es proporcional a la cantidad de pigmentos fotosintéticos que presentan las plantas. Ya comienzan a evidenciarse diferencias entre la distribución pigmentaria de las especies de hoja verde y las de hoja roja-granate, cuya comparación se realizará en el apartado de procesamiento de datos.
3.3. Tasas de fotosíntesis
Cada uno de los productos de la fotosíntesis es liberado o sintetizado en cada una de las fases en las que puede dividirse el proceso. Los pigmentos fotosintéticos participan en la fase dependiente de la luz, en la que se libera también el oxígeno. Por esta razón se basará la medida de tasa de fotosíntesis en la cantidad de oxígeno producido por las plantas en el periodo de medición.
Para medir esta variable se utilizó un sensor específico de concentración de oxígeno que usa una celda electroquímica para su medición. La celda contiene un ánodo de plomo y un cátodo de oro inmersos en un electrolito, de forma que las moléculas de oxígeno que entran en la celda son electroquímicamente reducidas. Esta reacción genera una corriente de electrones que es proporcional a la concentración de oxígeno entre los electrodos, registrándose de este modo el % de oxígeno en el recinto cerrado en el que se encuentran las plantas10.
La alta sensibilidad del sensor de oxígeno determinaba que para conseguir unas medidas fiables las plantas debían encontrarse en un recinto aislado y sin incidencia directa de la luz del sol. Para cumplir estas condiciones se planteaba la necesidad de idear un montaje específico. Primeramente se optó por introducir las plantas en bombonas de plástico transparente, pero éstas no eran lo suficientemente grandes para introducir también la maceta de las plantas; se consideró que el montaje no era adecuado ya que el oxígeno podría escaparse con facilidad. Finalmente, se construyó una estructura de madera con forma octogonal con un soporte en la parte superior en el que poder fijar el sensor de registro de concentración de oxígeno. La estructura estaba rodeada con un plástico traslúcido lo suficientemente amplio para rodear la planta por completo, de esta forma se protegía el sensor de la acción directa del sol y la planta se mantenía completamente aislada.

El Instituto Rosa Chacel, dónde se realizaron estas pruebas, consta cono un invernadero. Sin embargo, se consideró que no era adecuado realizarlas en este lugar, ya que la protección del tejado del invernadero unida al plástico traslúcido del montaje suponía una excesiva limitación de la luminosidad. El método seguido fue:
- Las plantas presentaban diferente porte, por lo que antes de realizar las mediciones se podaron para que todas tuvieran una superficie foliar aproximadamente igual.
- Se colocaron las plantas una a una en el interior de la campana y se cerró el plástico recogiéndolo debajo de la maceta de la planta.
- La monitorización del proceso de fotosíntesis se realizó una sola vez con cada planta, durante 15 minutos y tomando valores en un rango de cuatro de segundos.
A continuación se muestra el registro de la tasa de fotosíntesis con respecto al tiempo del grupo de plantas con hoja verde y de plantas con hoja roja-granate. Se ha realizado la media del registro individual de cada una de las plantas para conseguir una distribución media de ambos grupos.
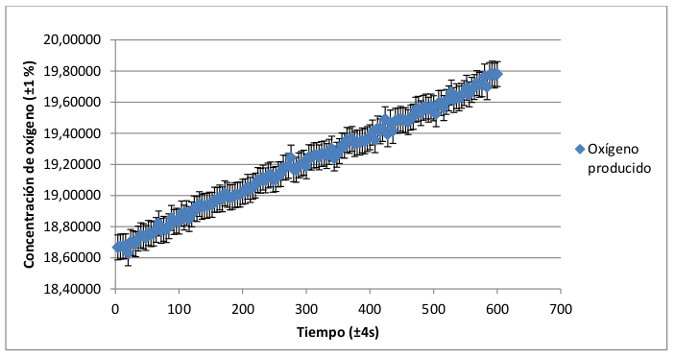
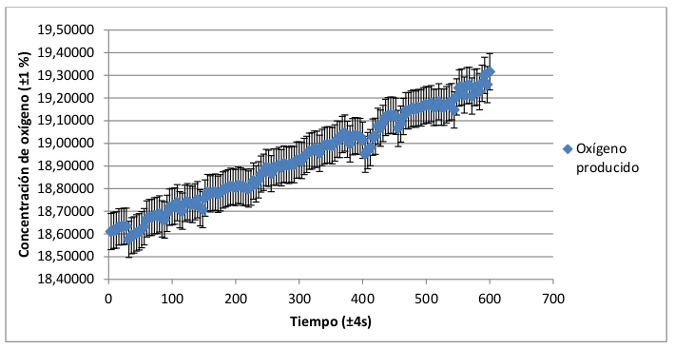
Aparentemente la distribución que describen los datos registrados para ambos grupos de plantas es muy similar, por lo que es necesario realizar un análisis comparativo de ambas, que se relacionará en el apartado de conclusiones con el análisis de la distribución pigmentaria de plantas de hoja verde y plantas de hoja roja-granate.
3.4. Procesamiento de datos
a. Comparación de la distribución pigmentaria
Primeramente se ha realizado la media de las distribuciones pigmentarias de por un lado las plantas de hojas verdes y por otro las plantas de hojas rojas-granates.
Se debe tener en cuenta que la especie Chlorophytum comosum no interviene en la media de las plantas de hojas verdes, puesto que los resultados de sus cromatografías son muy diferentes al patrón que siguen en resto de plantas de este grupo. De hecho, su composición pigmentaria es más similar a la de las plantas de hojas rojas-granates, por lo que se ha considerado que puede prescindirse de sus datos ya que podrían influir en las conclusiones posteriores.

Se observa una clara diferencia en la cantidad de clorofilas y carotenos, de colores verde y rojizo respectivamente. Las plantas de hoja verde efectivamente tienen mayor concentración en clorofila y las plantas de hoja roja-granate mayor concentración de carotenos. Estos dos pigmentos corresponden aproximadamente al 60% de la composición pigmentaria total de ambos grupos, lo que corrobora la hipótesis de partida: el color de las hojas de las plantas se corresponde con el color su pigmento fotosintético mayoritario.
Por otro lado, las plantas de hoja roja-granate presentan mayor concentración de clorofilas (28% aprox.) que el que presentan las plantas de hoja verde de carotenos (23% aprox.). Esto corrobora el papel importante de la clorofila en los fotosistemas de los cloroplastos de las plantas.
Finalmente las plantas de hoja verde son las que presentan mayor concentración de xantofilas, incluso mayor de la que presenta de caroteno, de lo que puede deducirse cierta superioridad de las xantofilas sobre los carotenos en el papel de pigmentos accesorios. Sin embargo las xantofilas no se han analizado tan específicamente como los carotenos o las clorofilas.
b. Análisis de tasas de fotosíntesis
El registro de la tasa de producción de oxígeno de plantas de hoja verde y de hoja roja-granate presenta un claro crecimiento en función del tiempo. Para conocer más acerca de la dependencia de las variables concentración de oxígeno y tiempo se ha calculado la recta de regresión lineal a la que mejor se ajustan los datos registrados. Del mismo modo se ha calculado el coeficiente de correlación de Pearson referida a cada una de ellas, que indicará el grado de ajuste de los datos a la distribución lineal.
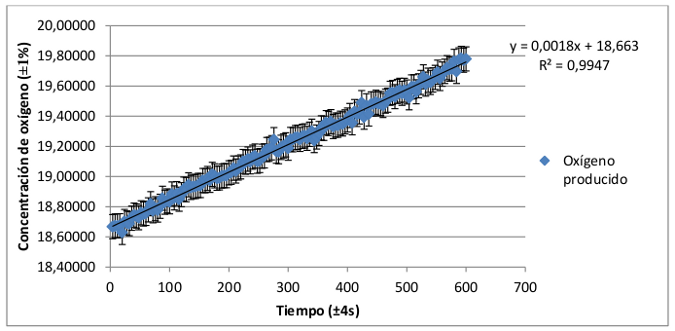
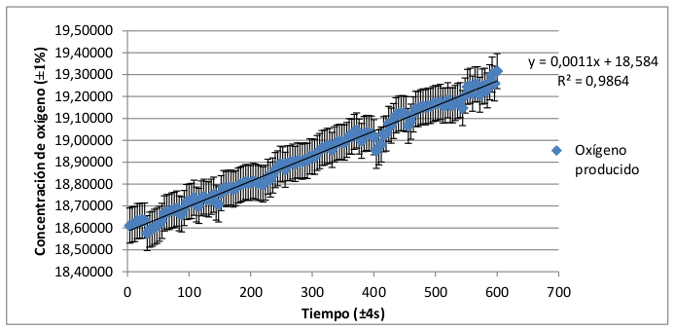
Dado que los coeficientes de correlación múltiple de Pearson (R2) son muy próximos a 1, (0,9947 para el grupo de plantas de hoja verde y 0,9864 para el de hoja roja-granate) puede determinarse que la producción de oxígeno en las plantas angiospermas aumenta según una distribución lineal con respecto al tiempo.
De esta forma, una mayor pendiente en esta distribución lineal indicará una mayor producción de oxígeno y con ello mayor eficacia en el proceso de fotosíntesis. La diferencia de las pendientes de la distribución de la producción de oxígeno de ambos grupos es mínima, aproximadamente el 0,0007. De ello puede concluirse que no existen diferencias significativas entre ambas tasas de producción. Por tanto, las plantas angiospermas, independientemente de su composición pigmentaria, desarrollan un proceso de fotosíntesis igualmente eficaz.
4. Conclusiones
A la luz de los resultados obtenidos en la investigación se ha establecido que no existe diferencia significativa entre la tasa de fotosíntesis de plantas de hoja verde y plantas de hoja roja-granate; aunque sí se diferencian en su composición pigmentaria. Se concluye que el desarrollo de las plantas angiospermas no depende del color de sus hojas, es decir, del pigmento mayoritario que presentan, aunque estos participen activamente en la fase lumínica de la fotosíntesis, precursora del proceso.
La clorofila es el pigmento mayoritario en las plantas de hoja verde y su rango de absorción se encuentra en los 450 nm y los 670 nm de longitud de onda, zonas de máxima energía. Sin embargo, las plantas de hoja roja-granate, en las que la cantidad de clorofila es menor que la cantidad de carotenos, alcanzan prácticamente el mismo rendimiento en el proceso de fotosíntesis. Se deduce que los pigmentos fotosintéticos accesorios (carotenos, en este caso) suplen las necesidades de la planta en aquellas especies en las que la cantidad de clorofila no es suficiente. Aumentan el rango de longitud de onda que absorbe la planta y por tanto la tasa de fotosíntesis.
Las conclusiones de la investigación han dado lugar a nuevos interrogantes y al planteamiento de nuevos problemas con los que podría darse continuidad al estudio. Se propone cultivar una planta con alta concentración de clorofilas, por ejemplo Impatiens wallwriana, en un medio sin magnesio, componente esencial de la molécula de clorofila y comprobar cómo respondería la planta, ¿desarrollaría otros pigmentos accesorios y cambiaría su coloración externa?
En definitiva, a pesar de las limitaciones consideradas en esta investigación, como haber utilizado un método muy sencillo para medir los pigmentos fotosintéticos de las plantas en vez de técnicas más precisas como la espectroscopia; este estudio ha servido para descartar la hipótesis de partida. Se ha concluido que el éxito adaptativo que presentan las plantas de hoja verde sobre las plantas de hoja roja-granate no se debe a una mayor eficacia en el desarrollo de la fotosíntesis. De este modo queda abierto un nuevo interrogante, ¿a qué se debe entonces la evolución favorable de las plantas de hoja verde frente a las de hoja roja-granate?
5. Bibliografía
- BERG, Linda. Introductory Botany: Plants, People, and the Environment. Segunda Edición. Thomson Higher Education. Belmont, USA. 2008. IBN: 978-0534-46669-5
- BLANCO, Inmaculada; GRANADO, Fernando; OLMEDILLA, Begoña. Carotenoides y salud humana. Fundación Española de la Nutrición (F.E.N). “Serie Informes”. Unidad de Vitaminas. Sección de Nutrición. Clínica Puerta de Hierro, Madrid, España. 2001. ISBN: 84-930544-2-9
- BLANKENDHIP, Robert E. Mollecular mechanisms of photosynthesis. Blackwell Publishing.Williston, USA. 2002. ISBN: 0-632-04321-0
- COLLAZO ORTEGA, Margarita; RODÉS GARCÍA, Rosa. Manual de prácticas de fotosíntesis. Primera Edición. Universidad Nacional Autónoma de México. Facultad de Ciencias. México, 2006. ISBN: 970-32-3313-9
- GARCÍA, C. Ecología Forestal: Estructura, Funcionamiento y Producción de las masas forestales. (Apartado 6: Fotosíntesis)
- MANRIQUE REOL, Esteban. Ecosistemas. Revista científica y técnica de ecología y medio ambiente. Madrid, España. 2003.
- NEIL, A. Campbell, Jane B. Biología. Séptima Edición. Editorial Médica Panamericana. Madrid, España. 2007. ISBN: 978-84-7903-998-1
- STARR, Cecie; TAGGART, Ralph. Biología. La unidad y la diversidad de la vida. Undécima Edición. Cengage Learning Editores. Santa Fe, México, D.F. 2008. ISBN: 10-970-686-777-5
- TAIZ, Linciln; ZEIGER, Eduardo. Fisiología vegetal. Publicaciones de la Universidad Jaime I. Barcelona, España. 2002. ISBN: 978-84-8027-601-2
- http://tsdb27.blogspot.com/2011/06/fotosintesis.html (Consultado el 16 de Febrero de 2012)
- http://es.wikipedia.org/wiki/Archivo:Chlorophyll_structure.png (Consultado el 26 de Febrero de 2012)
6. Anexo I. Imágenes


1. BLANKENSHIP, Robert E. Molecular mechanism of photosynthesis. Ed. Blackwell Publishing. Página 4.↑
2. NEIL A. Campbell, Jane B. Biología. Ed. Panamericana. Página 186.↑
3. STARR, Cecie; TAGGART, Ralph. Biología. La unidad y la diversidad de la vida. Ed. Cengage Learning. Pág. 110.↑
4. BERG, Linda. Introductory Botany: Plants, People, and the Environment. Thomson Higher Education. Página 72.↑
5. http://es.wikipedia.org/wiki/Archivo:Chlorophyll_structure.png (Consultado el 26 de Febrero de 2012).↑
6. BLANCO, Inmaculada; GRANADO, Fernando; OLMEDILLA, Begoña. Carotenoides y salud humana. Página 13.↑
7. MANRIQUE REOL, Esteban. Ecosistemas. Revista científica y técnica de ecología y medio ambiente. Página 5.↑
8. http://tsdb27.blogspot.com/2011/06/fotosintesis.html (Actualizado 23-Junio-2011; consultado 16-Febrero-2012).↑
9. COLLAZO ORTEGA, Margarita. GARCÍA ROLDÉS, Rosa. Manual de Prácticas de Fotosíntesis. Página 27.↑
10. Vernier LabQuest ® Mini. User’s Manual © 2010.↑